| |
Developmental biology (W) |
Developmental biology
Developmental biology (W)
|
| |
Perspectives
|
Perspectives
Perspectives (W)
The main processes involved in the embryonic development of animals are: regional specification, morphogenesis, cell differentiation, growth, and the overall control of timing explored in evolutionary developmental biology:
- Regional specification refers to the processes that create spatial pattern in a ball or sheet of initially similar cells. This generally involves the action of cytoplasmic determinants, located within parts of the fertilized egg, and of inductive signals emitted from signaling centers in the embryo. The early stages of regional specification do not generate functional differentiated cells, but cell populations committed to develop to a specific region or part of the organism. These are defined by the expression of specific combinations of transcription factors.
- Morphogenesis relates to the formation of three-dimensional shape. It mainly involves the orchestrated movements of cell sheets and of individual cells. Morphogenesis is important for creating the three germ layers of the early embryo (ectoderm, mesoderm and endoderm) and for building up complex structures during organ development.
- Cell differentiation relates specifically to the formation of functional cell types such as nerve, muscle, secretory epithelia etc. Differentiated cells contain large amounts of specific proteins associated with the cell function.
- Growth involves both an overall increase in size, and also the differential growth of parts (allometry) which contributes to morphogenesis. Growth mostly occurs through cell division but also through changes of cell size and the deposition of extracellular materials.
- The control of timing of events and the integration of the various processes with one another is the least well understood area of the subject. It remains unclear whether animal embryos contain a master clock mechanism or not.
The development of plants involves similar processes to that of animals. However plant cells are mostly immotile so morphogenesis is achieved by differential growth, without cell movements. Also, the inductive signals and the genes involved are different from those that control animal development. |

|
|
|
|
| |
Developmental processes
|
Cell differentiation
Cell differentiation (W)
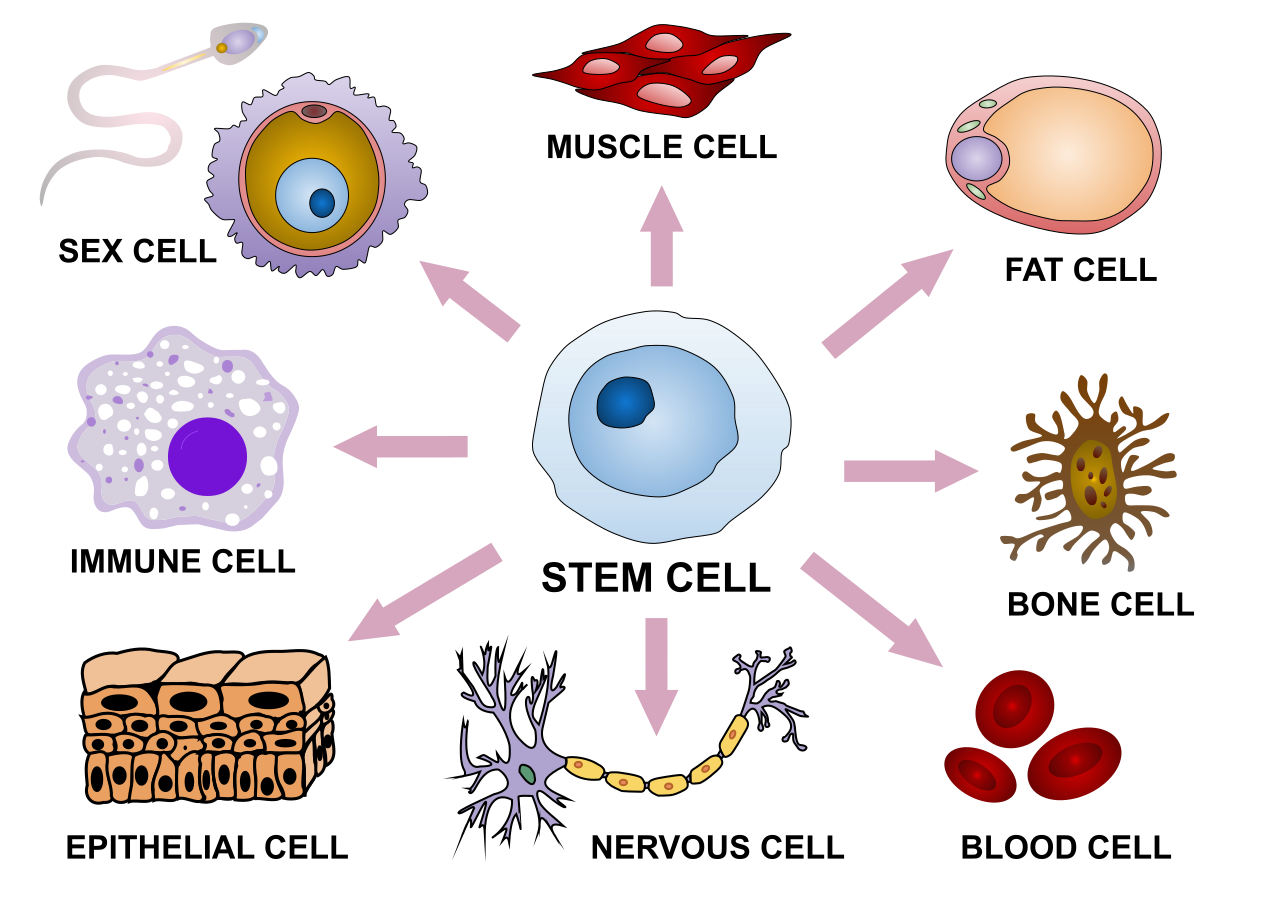
Cell differentiation is the process whereby different functional cell types arise in development. For example, neurons, muscle fibers and hepatocytes (liver cells) are well known types of differentiated cells. Differentiated cells usually produce large amounts of a few proteins that are required for their specific function and this gives them the characteristic appearance that enables them to be recognized under the light microscope. The genes encoding these proteins are highly active. Typically their chromatin structure is very open, allowing access for the transcription enzymes, and specific transcription factors bind to regulatory sequences in the DNA in order to activate gene expression. For example, NeuroD is a key transcription factor for neuronal differentiation, myogenin for muscle differentiation, and HNF4 for hepatocyte differentiation. Cell differentiation is usually the final stage of development, preceded by several states of commitment which are not visibly differentiated. A single tissue, formed from a single type of progenitor cell or stem cell, often consists of several differentiated cell types. Control of their formation involves a process of lateral inhibition, based on the properties of the Notch signaling pathway. For example, in the neural plate of the embryo this system operates to generate a population of neuronal precursor cells in which NeuroD is highly expressed.
.png)
The Notch-delta system in neurogenesis.(Slack Essential Dev Biol Fig 14.12a). (L) |
|
|
|
|
|
|
Regeneration
Regeneration (W)
Regeneration indicates the ability to regrow a missing part. This is very prevalent amongst plants, which show continuous growth, and also among colonial animals such as hydroids and ascidians. But most interest by developmental biologists has been shown in the regeneration of parts in free living animals. In particular four models have been the subject of much investigation. Two of these have the ability to regenerate whole bodies: Hydra, which can regenerate any part of the polyp from a small fragment, and planarian worms, which can usually regenerate both heads and tails. Both of these examples have continuous cell turnover fed by stem cells and, at least in planaria, at least some of the stem cells have been shown to be pluripotent. The other two models show only distal regeneration of appendages. These are the insect appendages, usually the legs of hemimetabolous insects such as the cricket, and the limbs of urodele amphibians. Considerable information is now available about amphibian limb regeneration and it is known that each cell type regenerates itself, except for connective tissues where there is considerable interconversion between cartilage, dermis and tendons. In terms of the pattern of structures, this is controlled by a re-activation of signals active in the embryo. There is still debate about the old question of whether regeneration is a "pristine" or an "adaptive" property. If the former is the case, with improved knowledge, we might expect to be able to improve regenerative ability in humans. If the latter, then each instance of regeneration is presumed to have arisen by natural selection in circumstances particular to the species, so no general rules would be expected. |
|
|
|
|
| |
Embryonic development of animals
|
Embryonic development of animals
Embryonic development of animals (W)
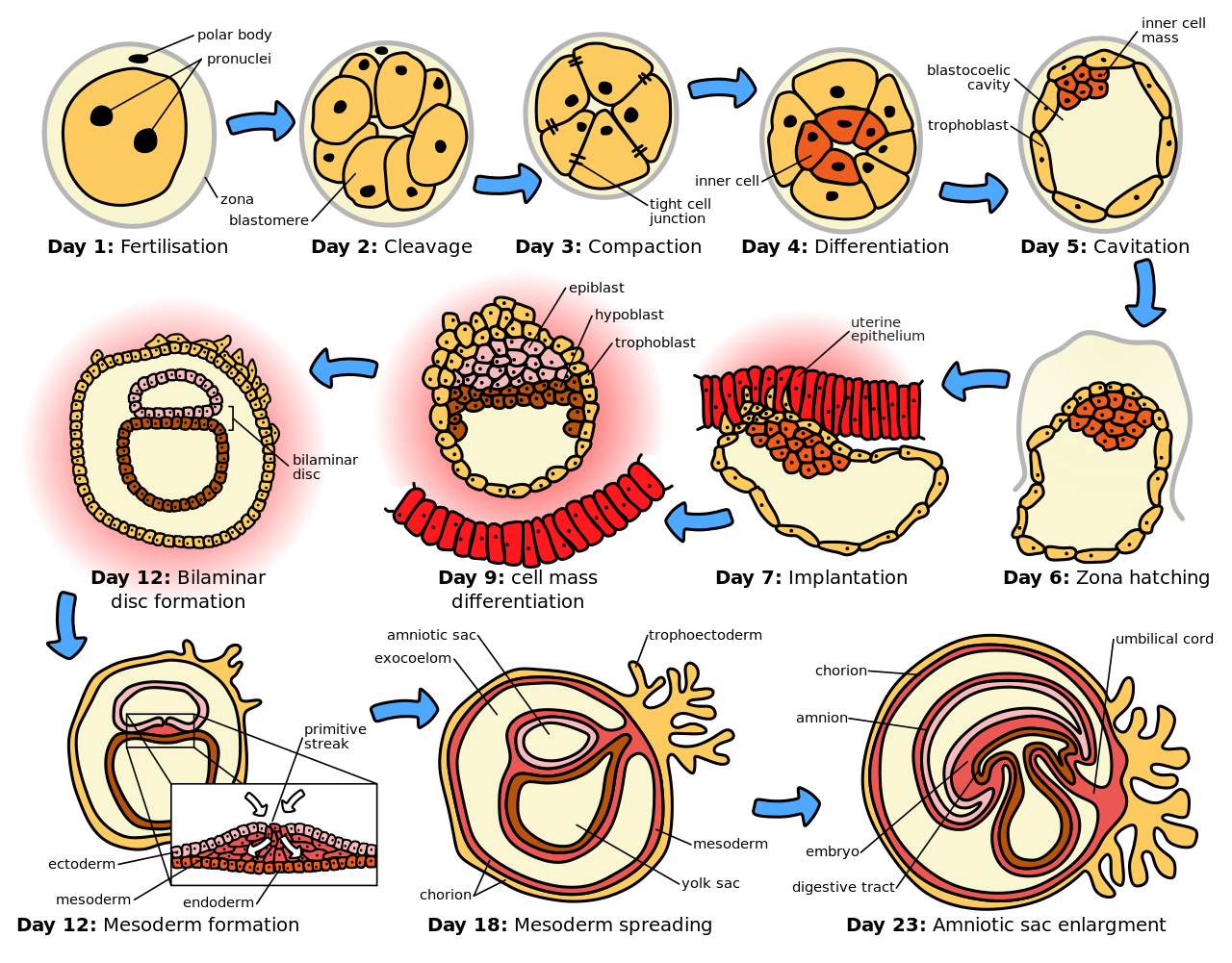
The sperm and egg fuse in the process of fertilization to form a fertilized egg, or zygote. This undergoes a period of divisions to form a ball or sheet of similar cells called a blastula or blastoderm. These cell divisions are usually rapid with no growth so the daughter cells are half the size of the mother cell and the whole embryo stays about the same size. They are called cleavage divisions.
Mouse epiblast primordial germ cells (see Figure: “The initial stages of human embryogenesis”) undergo extensive epigenetic reprogramming. This process involves genome-wide DNA demethylation, chromatin reorganization and epigenetic imprint erasure leading to totipotency. DNA demethylation is carried out by a process that utilizes the DNA base excision repair pathway.
Morphogenetic movements convert the cell mass into a three layered structure consisting of multicellular sheets called ectoderm, mesoderm and endoderm. These sheets are known as germ layers. This is the process of gastrulation. During cleavage and gastrulation the first regional specification events occur. In addition to the formation of the three germ layers themselves, these often generate extraembryonic structures, such as the mammalian placenta, needed for support and nutrition of the embryo, and also establish differences of commitment along the anteroposterior axis (head, trunk and tail).
Regional specification is initiated by the presence of cytoplasmic determinants in one part of the zygote. The cells that contain the determinant become a signaling center and emit an inducing factor. Because the inducing factor is produced in one place, diffuses away, and decays, it forms a concentration gradient, high near the source cells and low further away. The remaining cells of the embryo, which do not contain the determinant, are competent to respond to different concentrations by upregulating specific developmental control genes. This results in a series of zones becoming set up, arranged at progressively greater distance from the signaling center. In each zone a different combination of developmental control genes is upregulated. These genes encode transcription factors which upregulate new combinations of gene activity in each region. Among other functions, these transcription factors control expression of genes conferring specific adhesive and motility properties on the cells in which they are active. Because of these different morphogenetic properties, the cells of each germ layer move to form sheets such that the ectoderm ends up on the outside, mesoderm in the middle, and endoderm on the inside. Morphogenetic movements not only change the shape and structure of the embryo, but by bringing cell sheets into new spatial relationships they also make possible new phases of signaling and response between them.
Growth in embryos is mostly autonomous. For each territory of cells the growth rate is controlled by the combination of genes that are active. Free-living embryos do not grow in mass as they have no external food supply. But embryos fed by a placenta or extraembryonic yolk supply can grow very fast, and changes to relative growth rate between parts in these organisms help to produce the final overall anatomy.
The whole process needs to be coordinated in time and how this is controlled is not understood. There may be a master clock able to communicate with all parts of the embryo that controls the course of events, or timing may depend simply on local causal sequences of events.
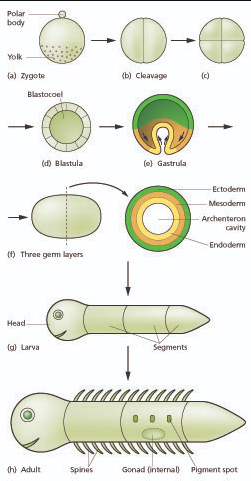
Generalized scheme of embryonic development. Slack "Essential Developmental Biology" Fig.2.8
____________. |
|
|
|
|
|
|
Metamorphosis
Metamorphosis (W)
Developmental processes are very evident during the process of metamorphosis. This occurs in various types of animal. Well-known examples are seen in frogs, which usually hatch as a tadpole and metamorphoses to an adult frog, and certain insects which hatch as a larva and then become remodeled to the adult form during a pupal stage.
All the developmental processes listed above occur during metamorphosis. Examples that have been especially well studied include tail loss and other changes in the tadpole of the frog Xenopus, and the biology of the imaginal discs, which generate the adult body parts of the fly Drosophila melanogaster. |
|
|
|
| |
Plant development
|
Plant development
Plant development (W)
Plant development is the process by which structures originate and mature as a plant grows. It is studied in plant anatomy and plant physiology as well as plant morphology.
Plants constantly produce new tissues and structures throughout their life from meristems located at the tips of organs, or between mature tissues. Thus, a living plant always has embryonic tissues. By contrast, an animal embryo will very early produce all of the body parts that it will ever have in its life. When the animal is born (or hatches from its egg), it has all its body parts and from that point will only grow larger and more mature.
The properties of organization seen in a plant are emergent properties which are more than the sum of the individual parts. "The assembly of these tissues and functions into an integrated multicellular organism yields not only the characteristics of the separate parts and processes but also quite a new set of characteristics which would not have been predictable on the basis of examination of the separate parts." |
|
|
|
|
Growth
Growth (W)
A vascular plant begins from a single celled zygote, formed by fertilisation of an egg cell by a sperm cell. From that point, it begins to divide to form a plant embryo through the process of embryogenesis. As this happens, the resulting cells will organize so that one end becomes the first root, while the other end forms the tip of the shoot. In seed plants, the embryo will develop one or more "seed leaves" (cotyledons). By the end of embryogenesis, the young plant will have all the parts necessary to begin its life.
Once the embryo germinates from its seed or parent plant, it begins to produce additional organs (leaves, stems, and roots) through the process of organogenesis. New roots grow from root meristems located at the tip of the root, and new stems and leaves grow from shoot meristems located at the tip of the shoot. Branching occurs when small clumps of cells left behind by the meristem, and which have not yet undergone cellular differentiation to form a specialized tissue, begin to grow as the tip of a new root or shoot. Growth from any such meristem at the tip of a root or shoot is termed primary growth and results in the lengthening of that root or shoot. Secondary growth results in widening of a root or shoot from divisions of cells in a cambium.
In addition to growth by cell division, a plant may grow through cell elongation. This occurs when individual cells or groups of cells grow longer. Not all plant cells will grow to the same length. When cells on one side of a stem grow longer and faster than cells on the other side, the stem will bend to the side of the slower growing cells as a result. This directional growth can occur via a plant's response to a particular stimulus, such as light (phototropism), gravity (gravitropism), water, (hydrotropism), and physical contact (thigmotropism).
Plant growth and development are mediated by specific plant hormones and plant growth regulators (PGRs) (Ross et al. 1983). Endogenous hormone levels are influenced by plant age, cold hardiness, dormancy, and other metabolic conditions; photoperiod, drought, temperature, and other external environmental conditions; and exogenous sources of PGRs, e.g., externally applied and of rhizospheric origin. |
|
|
|
|
Morphological variation
Morphological variation (W)
Plants exhibit natural variation in their form and structure. While all organisms vary from individual to individual, plants exhibit an additional type of variation. Within a single individual, parts are repeated which may differ in form and structure from other similar parts. This variation is most easily seen in the leaves of a plant, though other organs such as stems and flowers may show similar variation. There are three primary causes of this variation: positional effects, environmental effects, and juvenility. |
|
|
|
Evolution of plant morphology
Evolution of plant morphology (W)
Transcription factors and transcriptional regulatory networks play key roles in plant morphogenesis and their evolution. During plant landing, many novel transcription factor families emerged and are preferentially wired into the networks of multicellular development, reproduction, and organ development, contributing to more complex morphogenesis of land plants.
Most land plants share a common ancestor, multicellular algae. An example of the evolution of plant morphology is seen in charophytes. Studies have shown that charophytes have traits that are homologous to land plants. There are two main theories of the evolution of plant morphology, these theories are the homologous theory and the antithetic theory. The commonly accepted theory for the evolution of plant morphology is the antithetic theory. The antithetic theory states that the multiple mitotic divisions that take place before meiosis, cause the development of the sporophyte. Then the sporophyte will development as an independent organism. |
|
|
|
|
| |
Developmental model organisms
|
Developmental model organisms
Developmental model organisms (W)
Much of developmental biology research in recent decades has focused on the use of a small number of model organisms. It has turned out that there is much conservation of developmental mechanisms across the animal kingdom. In early development different vertebrate species all use essentially the same inductive signals and the same genes encoding regional identity. Even invertebrates use a similar repertoire of signals and genes although the body parts formed are significantly different. Model organisms each have some particular experimental advantages which have enabled them to become popular among researchers. In one sense they are "models" for the whole animal kingdom, and in another sense they are "models" for human development, which is difficult to study directly for both ethical and practical reasons. Model organisms have been most useful for elucidating the broad nature of developmental mechanisms. The more detail is sought, the more they differ from each other and from humans.
Plants:
Vertebrates:
- Frog: Xenopus (X.laevis and tropicalis). Good embryo supply. Especially suitable for microsurgery.
- Zebrafish: Danio rerio. Good embryo supply. Well developed genetics.
- Chicken: Gallus gallus. Early stages similar to mammal, but microsurgery easier. Low cost.
- Mouse: Mus musculus. A mammal with well developed genetics.
Invertebrates:
Also popular for some purposes have been sea urchins and ascidians. For studies of regeneration urodele amphibians such as the axolotl Ambystoma mexicanum are used, and also planarian worms such as Schmidtea mediterranea. Organoids have also been demonstrated as an efficient model for development. Plant development has focused on the thale cress Arabidopsis thaliana as a model organism. |
|
|
|
|
|
|
|
|
|
|
| |
Biological development (B) |
Biological development (B)
Biological development (B)
Introduction (W)
|
| |
Biological development, the progressive changes in size, shape, and function during the life of an organism by which its genetic potentials (genotype) are translated into functioning mature systems (phenotype). Most modern philosophical outlooks would consider that development of some kind or other characterizes all things, (?) in both the physical and biological worlds. Such points of view go back to the very earliest days of philosophy.
Among the pre-Socratic philosophers of Greek Ionia, half a millennium before Christ, some, like Heracleitus, believed that all natural things are constantly changing. In contrast, others, of whom Democritus is perhaps the prime example, suggested that the world is made up by the changing combinations of atoms, which themselves remain unaltered, not subject to change or development. The early period of post-Renaissance European science may be regarded as dominated by this latter atomistic view, which reached its fullest development in the period between Newton’s laws of physics and Dalton’s atomic theory of chemistry in the early 19th century. This outlook was never easily reconciled with the observations of biologists, and in the last hundred years a series of discoveries in the physical sciences have combined to swing opinion back toward the Heracleitan emphasis on the importance of process and development. {= becoming?} The atom, which seemed so unalterable to Dalton, has proved to be divisible after all, and to maintain its identity only by processes of interaction between a number of component subatomic particles, which themselves must in certain aspects be regarded as processes rather than matter. Albert Einstein’s theory of relativity showed that time and space are united in continuum, which implies that all things are involved in time; that is to say, in development. {!}
The philosophers who charted the transition from the nondevelopmental view, for which time was an accidental and inessential element, were Henri Bergson and, in particular, Alfred North Whitehead. Karl Marx and Friedrich Engels, with their insistence on the difference between dialectical and mechanical materialism, may be regarded as other important innovators of this trend, {?} although the generality of their philosophy was somewhat compromised by the political context in which it was placed and the rigidity with which their later followers have interpreted it. {?}
Philosophies of the Heracleitan type, which emphasize process and development, provide much more appropriate frameworks for biology than do philosophies of the atomistic kind. Living organisms confront biologists with changes of various kinds, all of which could be regarded as in some sense developmental; however, biologists have found it convenient to distinguish the changes and to use the word development for only one of them. Biological development can be defined as the series of progressive, nonrepetitive changes that occur during the life history of an organism. The kernel of this definition is to contrast development with, on the one hand, the essentially repetitive chemical changes involved in the maintenance of the body, which constitute “ metabolism,” and on the other hand, with the longer term changes, which, while nonrepetitive, involve the sequence of several or many life histories, and which constitute evolution.
As with most formal definitions, these distinctions cannot always be applied strictly to the real world. In the viruses, for instance, and even in bacteria, it is difficult to make a distinction between metabolism and development, since the metabolic activity of a virus particle consists of little more than the development of new virus particles. In certain other cases, the distinction between development and evolution becomes blurred: the concept of an individual organism with a definite life history may be very difficult to apply in plants that reproduce by vegetative division, the breaking off of a part that can grow into another complete plant. The possibilities for debate that arise in these special cases, however, do not in any way invalidate the general usefulness of the distinctions as conventionally made in biology. |
|
| |
| |
The Scope Of Development
|
The Scope Of Development
The Scope Of Development (B)
All organisms, including the very simplest, consist of two components, distinguished by a German biologist, August Weismann, at the end of the 19th century, as the “ germ plasm” and the “ soma.” The germ plasm consists of the essential elements, or genes, passed on from one generation to the next, and the soma consists of the body that may be produced as the organism develops. In more modern terms, Weismann’s germ plasm is identified with DNA ( deoxyribonucleic acid), which carries, encoded in the complex structure of its molecule, the instructions necessary for the synthesis of the other compounds of the organism and their assembly into the appropriate structures. It is this whole collection of other compounds (proteins, fats, carbohydrates, and others) and their arrangement as a metabolically functioning organism that constitutes the soma. Biological development encompasses, therefore, all the processes concerned with implementing the instructions contained in the DNA. Those instructions can only be carried out by an appropriate executive machinery, the first phase of which is provided by the cell that carries the DNA into the next generation: in animals and plants by the fertilized egg cell; in viruses by the cell infected. In life histories that have more than a minimal degree of complexity, the executive machinery itself becomes modified as the genetic instructions are gradually put into operation, and new mechanisms of protein synthesis are brought into functional condition. The fundamental problem of developmental biology is to understand the interplay between the genetic instructions and the mechanisms by which those instructions are carried out.
In the language of genetics the word genotype is used to indicate the hereditary instructions passed on from one generation to another in the genes, while phenotype is the term given to the functioning organisms produced by those instructions. Biological development, therefore, consists of the production of phenotypes. The point made in the last paragraph is that the formation of the phenotype of one generation depends on the functioning of part of the phenotype of the previous generation (e.g., egg cell), as the mechanism that begins the interpretation of the instructions contained in the new organism’s genotype. |
|
|
|
|
Types of development
Types of development (B)
In the entire realm of organisms, many different modes of development are found, the most important categories of which can be discussed as pairs of contrasting types |
|
|
|
Quantitative and qualitative development
Quantitative and qualitative development (B)
Development may amount to no more than a quantitative change (usually an increase) in a system that remains essentially unaltered. Qualitative development involves an alteration in the nature of the system. Pure examples of the first type are difficult to find. Approximations to it occur when an animal or plant has attained a structure with the full complement of organs; it then appears to increase only in size, that is to say, quantitatively. This would be a period of simple growth. A closer examination nearly always shows that the system is also undergoing some qualitative change, however. A human infant at birth, for example, already has its full complement of organs, but the ensuing developmental period up to adulthood involves not only growth but also processes of maturation that involve qualitative as well as quantitative changes. Perhaps the most uncomplicated examples of quantitative development occur in certain simple plants and animals. Flatworms, for example, may become reduced in size when starved but increase in size again when provided with suitable nutrition; they thus undergo quantitative changes. Even in these cases, however, it is found that the constituent organs do not always merely become reduced in size but may actually suffer the loss of certain parts. |
|
|
|
|
Progressive and regressive development
Progressive and regressive development (B)
The normal processes of development in the majority of plants and animals may be considered progressive since they lead to increases in size and complexity and to the addition of new elements to the system. As already indicated, some organisms, when placed in adverse conditions, may undergo regressive changes, both in size and complexity. Such regressive changes are a part of the normal life history of certain organisms. Characteristically, these are species in which the organism at an early stage develops a relatively complex structure that enables it to be motile, and later adopts a form of life for which motility is no longer a necessity. A good example is that of the barnacles, a group of marine crustaceans in which the egg at first develops into a motile larva that soon settles down and becomes firmly attached to a solid underwater surface. The barnacle then loses many of the organs characteristic of the motile phase and develops into its familiar stationary form.
There are a number of other examples, particularly in groups in which the adults adopt a parasitic form of life, especially within the digestive system or other tissues of a host animal, from which they have only to absorb their nutriment without having to move or to possess suitable organs for capturing prey. In such cases the early developmental period is characterized by progression toward more complex forms followed by a period of regression in which many of these organs may be lost. During this regressive period certain components of the organism (i.e., those concerned with functioning as a sessile or parasitic form) may undergo progressive development at the same time as the other organs are regressing. |
|
|
|
|
Single-phase and multiphase development
Single-phase and multiphase development (B)
The most familiar organisms, including man, undergo a single-phase development; the organs that appear at early stages persist throughout the whole of life. There are many kinds of animals that develop one or more larval stages adapted to a life different from that of the adult. Perhaps the best known of these is the common frog. The egg first develops into a tadpole, which is provided with a large muscular tail by which it swims. The tadpole eventually undergoes a change of form, or metamorphosis. This involves the regression and resorption of the tail and the growth of the limbs. During this time the rest of the body of the tadpole undergoes less profound changes; the organs persist but undergo relatively far-reaching progressive changes. In other animals, the alteration between the larval and the adult forms may be much more drastic. The egg of a sea urchin, for instance, at first develops to a small larva (the pluteus), which is completely unlike that of the adult. During metamorphosis nearly all the structures of the pluteus disappear; the five-rayed adult develops from a very small rudiment within the larva. In other groups of marine invertebrates, there may be successive larval stages before the adult form appears.
Plants in general appear to exhibit a type of development related in a general way to the multiphased development just discussed in animals, although rather different from it in essence. This is called the “ alternation of generations.” The majority of higher plants possess two sets of similar chromosomes in each of their cells, that is to say they are diploid (2n), as are most higher animals. But in sexual reproduction, diploid cells undergo a reduction division so as to form precursors of the sex cells, which are haploid—i.e., they contain only one set of chromosomes. In animals these cells develop directly into the sex cells—egg and sperm—which unite in fertilization. In plants the haploid cells undergo some developmental processes before the functioning sex cells are produced. The products of this development are spoken of as the “haploid generation.” In most higher plants the haploid development is quite reduced, so that the haploid individuals contain only a few nuclei—those associated with the pollen tube on the male side and a few associated with the egg on the female side. In some lower plants, however, such as mosses and ferns, the haploid development may be much more extensive and give rise to quite sizable separate plants. In such cases a species contains two kinds of individuals, produced by different types of developmental processes controlled, however, by the same genotype. This may be compared with the multiphasing development of larval forms in animals. The situation in plants, however, is characterized by the two forms of the organism having different chromosomal constitutions—haploid and diploid—whereas the larval forms and the adult of an animal species have the same chromosomal constitution.
|
|
|
|
|
Structural and functional development
Structural and functional development (B)
These two categories cannot be regarded as a pair of opposites as were the previous pairs in this list; rather, they are two aspects of all processes of biological development and can be separated only conceptually, and for purposes of convenience of description. Function is the capacity of the biological system to carry out operations. At the level of the organism, these operations include walking, swimming, eating, digesting, etc.; at the cell level, typical functions are respiring, contracting, conducting nervous impulses, secreting hormones, etc.; and at the molecular level, all functions depend on the production of enzymes, coded by particular genes. Structure encompasses all parts of the organism capable of carrying out functions localized within the body of the organism and arranged in some particular spatial pattern. Contractile cells, for example, are grouped together to form muscle, and other cells are grouped together to form elements of the skeleton; both the muscles and the skeletal elements have definite spatial relations to each other.
These two aspects of development—function and structure—are not opposed to each other in any way. On the contrary, it is obvious that the higher level functions are clearly dependent on the proper structural relations and functions of cell systems. Even at the basic cellular or molecular levels, secretion or nervous conduction essentially depends upon the proper structural relation of the subcellular elements. It is, however, often convenient to focus discussion on one or other of these two aspects of development; for instance, a study may be made into the developmental processes that bring about the production of hemoglobin or insulin by a certain kind of cell, without at the moment being concerned with structural problems. Or again, the focus may be on the results of a certain process by which a mass of cells develops into a typical hand with five digits. In such an inquiry the structural aspects are paramount. |
|
|
|
|
Normal and abnormal development
Normal and abnormal development (B)
If a number of fertilized eggs of a given species are provided with conditions that enable them to develop at all, they will, with extraordinary regularity, develop into exceedingly similar adult organisms. The range of conditions they can tolerate is rather wide, and the similarity of the end products surprisingly complete. There are, indeed, good grounds for recognizing what must be considered normal development. The situation is perhaps more marked in animals than in plants, since the plants produced from a given batch of seed under a variety of environmental conditions often present considerably greater variation than is commonly found among animals. Even among plants, however, the differences produced by different conditions of cultivation are usually no more than quantitative differences in size and number of such organs as leaves and flowers, so that an individual can be described as well or poorly developed rather than as normally or abnormally developed. It is only in relatively few cases that a plant develops in quite different ways under two different conditions, neither of which can be considered abnormal or normal. In certain aquatic plants, for instance, the shape of the emergent leaves is different from the leaves that develop underwater. In such cases the plant actually has two normal forms of development.
It is possible, of course, to produce abnormal organisms by submitting a developing system to stimuli not usually encountered in a normal environment, such as certain chemicals. The presence of unusual genes also may result in deviations from the normal processes of development. In the vast majority of cases such abnormalities can be regarded as resulting from failure to carry out fully the normal processes of development. Functional abnormality in the adult consists in the failure of the system to produce a certain enzyme or functional cell type; a structural abnormality consists in the unusual appearance of certain component elements or in their arrangement in incompletely realized patterns. It is extremely rare to find examples in which the abnormality consists in the addition of a new enzyme not produced in normal development, or the formation of a new structural pattern of the elements.
One very important type of development that, from some points of view, can be considered as an exception to the rule that abnormal development is nearly always retrogressive, is carcinogenesis, the production of tumours. Carcinogenesis involves a change in the developmental behaviour of a group of cells. Initially, it often involves a loss of some of the functional and structural characteristics that previously appeared in the cells. It is commonly followed, however, by the assumption of new properties, which however untoward they may be for the host animal, must be considered as a progressive type: the cells often grow faster and multiply sooner than the noncancerous cells, for example. Furthermore, the cells may undergo a sequence of changes in character and in their arrangement within the tumour. All these features can be regarded in a developmental sense as progressive.
In view of the great rarity of cases of abnormal development that lead to progressive changes, it seems to follow that the organs produced during the normal development of any given species actually exhaust all the potentialities of its genotype for the production of orderly functional structures. It appears that the only abnormal developments that can be produced are either displacements of normal organs, or inadequacies in carrying out normal processes, or the initiation of progressive but quite disorderly processes, as in the production of tumours. |
|
|
|
|
General systems of development
General systems of development (B)
No text. |
|
|
|
Development of single-celled organisms
Development of single-celled organisms (B)
In viruses, activities consist in the production, aided by the machinery of a host cell, of units for building new virus or phage particles: development is simply the assemblage of these constituent units.
In the next higher grade of biological organization, the organism consists of a single cell. Many single-celled algae produce special forms of cells that correspond to the sex cells, or gametes; these cells may unite in fertilization, the resulting fertilized egg, or zygote, undergoing a short period of development. In many other single-celled organisms, however, reproduction takes place by the simple division of an original cell into two daughter cells. In such forms, development normally is part of the process of subdivision. It involves the remodelling of the parent cell into two smaller cells, which are then separated by the division. Something similar must, of course, be involved in the division of cells of higher organisms also. In many single-celled organisms, however, the cell contains a number of defined parts, which are arranged in very definite ways, so that the process of remodelling is very striking and easily observed. This is so, for instance, with ciliated protozoans, in which the cortex is provided with a large number of hairlike cilia or other appendages, arranged in precise patterns, and often with such other structures as a mouth or a gullet. These structures are reproduced in two identical but smaller copies during cell division. This does not necessarily imply that no other developmental processes are possible. The process of regeneration of parts removed occurs quite independently of cell division, for example. |
|
|
|
|
Open and closed systems of development
Open and closed systems of development (B)
There is a marked difference between the general system of development in multicellular plants and multicellular animals. In a plant, certain groups of cells retain throughout the whole life of the plant an embryonic capability to give rise to many types of cells. These regions, known as meristems, occur at the growing tips of branches and roots and as a cylindrical sheath around the stem. They consist of rapidly dividing cells capable of assembling into groups that form buds from which may arise new stems, leaves, flowers, or roots.
By contrast, most animals have no special regions that retain an embryonic character. In most forms, the whole egg, and the whole collection of cells immediately derived from it, take part in the developmental processes and form parts of the developing embryo. In some forms that go through a number of larval stages, the development of certain cells is interrupted at an early stage, and they are set aside and resume their development to form a later type of larva, or to form the adult after the larval stages are completed. An example would be the imaginal buds of some insects. The cells of these buds cannot be regarded as retaining a fully embryonic character comparable to that of the plant meristems, since they cannot perform all the developmental processes but only those involved in the production of the particular late-larval or adult structure for which they have been set aside. In general, then, plants remain embryonic in character, capable as it were of starting again from the beginning to carry out the entire developmental process. Their development is, in this sense, “open.” Most animals, on the other hand, lack persistently embryonic cells of this kind, and their development may be characterized as “closed.” (There may be certain exceptions to this in very simple forms, such as flatworms, in which certain cells called neoblasts seem able to participate in any type of development; these cells are usually scattered throughout the body, and the major developmental processes that bring into being the general form of the organism cannot be attributed to them, as the development of the plant can be attributed to the meristems.) |
|
|
|
|
Blastogenesis versus embryogenesis
Blastogenesis versus embryogenesis (B)
Some animals possess a second system of development, in contrast to the “closed” embryonic system emphasized in the last section. In its most fully developed form, this system consists in remodelling a portion of the parental body into a new organism without any involvement of eggs or sperm. In an adult hydra, a microscopic aquatic animal, a portion of the body may begin to grow exceptionally fast; its cells differentiate into the various cell types and become molded into the constituent organs to build up a new individual identical to the parent. The group of cells responsible for this behaviour is, in its early stages, referred to as a bud, or blastema. Before they become activated these cells may appear quite indistinguishable from the other cells of the body and betray no embryonic capability comparable to the meristems of plants.
In some higher organisms, including certain insects, reptiles, and amphibians, incomplete but still fairly extensive new developments of a similar kind may take place. They require the stimulus of an injury, however, which may involve the removal of part of the normal body. The usual result is a new development to regenerate, or replace, the missing part. The first stage in such regenerative processes consists in the formation of a blastema, that is, a group of rapidly dividing cells that shows little sign of cellular specialization. The evidence indicates that they may not arise, as was once thought, from persisting embryonic cells scattered within the adult body, but instead are formed of cells near the position of the injury. These cells lose their normal adult character and become capable of developing into most of the tissues required to replace the parts removed by the injury.
Development from a blastema, or blastogenesis, presents many contrasts to embryogenesis, the normal form of development from a fertilized egg. In blastogenesis, tissues that, during embryonic development, appear in sequence one after another, may be formed simultaneously and without any obvious sequential relations. Very little, however, is as yet understood about the mechanisms by which the various tissues within the blastema become differentiated from one another. It may well be that these mechanisms are more similar to those found in embryonic development than appears at first sight. |
|
|
|
|
Constituent processes of development
Constituent processes of development (B)
No text. |
|
|
|
Growth
Growth (B)
As was pointed out earlier, developing systems normally increase in size, at least during part of their development. “Growth” is a general term used to cover this phenomenon. It comprises two main aspects: (1) increase in cell numbers by cell division and (2) increase in cell size. These two processes may in some examples occur quite separately from each other; for instance, cells in certain rapidly growing tissues (e.g., the connective tissue or blood-forming systems in vertebrates) may increase greatly in number, while the cells remain approximately the same size. Alternatively, in some organs (e.g., the salivary glands of insects) the cells may increase greatly while remaining the same in number, each cell becoming enlarged, or hypertrophied. In such greatly enlarged cells there is often duplication of the genes, involving an increase in the DNA content of the nucleus, although no cell division takes place, and the nucleus continues as a single body, although with a multiplied, or “polyploid,” set of chromosomes.
In very many cases, however, the growth of an organ depends on increases both in cell number and in cell size. The relative importance of these two processes has yet to be properly investigated. One case that has been well studied is the size of the wings of the fruit fly Drosophila. The number of cells in the wing can be easily determined, since each bears a single hair that can be seen and counted in simple microscopic preparations. It has been found that there is an accommodation of factors: if there is an unusually large number of cells, these may be somewhat smaller than usual, so that the total size of the wing remains relatively unchanged.
Perhaps the major theoretical difficulty in the concept of growth is that it is a quantitative notion attached to an ill-defined entity. Growth is an increase in size; but size of what? If a cell or organ increases in volume merely by the absorption of water, or by the laying down of a mineral substance such as calcium carbonate, is this to be regarded as growth or not? |
|
|
|
|
Morphogenesis
Morphogenesis (B)
As was pointed out earlier, morphogenesis refers to all those processes by which parts of a developing system come to have a definite shape or to occupy particular relative positions in space. It may be regarded as the architecture of development. Morphogenetic processes involve the movement of parts of the developing system from one place to another in space, and therefore involve the action of physical forces, in contrast to processes of differentiation (see below), which require only chemical operations. Although in practice the physical and chemical processes of development normally proceed in close connection, for purposes of discussion it is often convenient to make an artificial separation between them.
There is an enormous variety of different kinds of structures within living organisms. They occur at all levels of size, from an elephant’s trunk to organelles within a cell, visible only with the electron microscope. There is still no satisfactory classification of the great range of processes by which these structures are brought into being. The following paragraphs constitute a tentative categorization that seems appropriate for the present state of biological thought on this topic.
Morphogenesis by differential growth
After their initiation, the various organs and regions of an organism may increase in size at different rates. Such processes of differential growth will change the overall shape of the body in which they occur. Processes of this kind take place very commonly in animals, particularly in the later stages of development. They are of major importance in the morphogenesis of plants, where the overall shape of the plant, the shape of individual leaves, and so on, depends primarily on the rates of growth of such component elements as the stems, the lateral shoots, and the vein and intervein material in leaves. In both animals and plants, such growth processes are greatly influenced by a variety of hormones. It is probable that factors internal to individual cells also always play a role.
Although differential growth may produce striking alterations in the general shape of organisms, these effects should probably be considered as somewhat superficial, since they only modify a basic pattern laid down by other processes. In a plant, for instance, the fundamental pattern is determined by the arrangement of the lateral buds around the central growing stem; whether these buds then grow fast or slowly relative to the stem is a secondary matter, however striking its results may be.
Morphogenetic fields
Many fundamental processes of pattern formation (e.g., the arrangement of lateral buds in growing plants) occur within areas or three-dimensional masses of tissue that show no obvious indications of where the various elements in the pattern will arise until they actually appear. Such masses of tissue, in which a pattern appears, have been spoken of as “fields.” This word was originally used in the early years of the 20th century by German authors who suggested an analogy between biological morphogenetic fields and such physical entities as magnetic or electromagnetic fields. The biological field is a description, but not an explanation, of the way in which the developing system behaves. The system develops as though each cell or subunit within it possessed “positional information” that specifies its location within the field and a set of instructions that lays down the developmental behaviour appropriate to each position.
There have been several attempts to account for the nature of the positional information and of the corresponding instructions. The oldest and best known of these is the gradient hypothesis. In many fields there is some region that is in some way “dominant,” so that the field appears as though organized around it. It is suggested that this region has a high concentration of some substance or activity, which falls off in a graded way throughout the rest of the field. The main deficiency of the hypothesis is that no one has yet succeeded in identifying satisfactorily the variables distributed in the gradients. Attempts to suppose that they are gradients of metabolic activity have, on investigation, always run into difficulties that can only be solved by defining metabolic activity in terms that reduce the hypothesis to a circular one in which metabolic activity is defined as that which is distributed in the gradient.
Recently, a new suggestion has been advanced concerning position information. Most processes within cells normally involve negative feedback control systems. These systems have a tendency to oscillate, or fluctuate regularly. In fact, any aspect of cell metabolism may be basically oscillatory in character; the cycle of cell growth and division may be only one example of a much more widespread phenomenon. The substances involved in these oscillations are likely to include diffusible molecules capable of influencing the behaviour of nearby cells. It is easy to envisage the possibility that there might be localized regions with oscillations of higher frequency or greater amplitude that act as centres from which trains of waves are radiated in all directions. It has been suggested that positional information is specified in terms of differences in phase between two or more such trains of transmitted oscillations.
Certain types of field phenomenon may involve an amplification of stochastic (random) variations. In systems containing a number of substances, with certain suitable rates of reaction and diffusion, chance variation on either side of an initial condition of equilibrium may become amplified both in amplitude and in the area involved. In this way, the processes may give rise to a pattern of differentiated areas, distributed in arrangements that depend on the boundary conditions.
Morphogenesis by the self-assembly of units
Complex structures may arise from the interaction between units that have characteristics such that they can fit together in a certain way. This is particularly appropriate for morphogenesis at the simple level of molecules or cells. Units such as the atoms of carbon, hydrogen, oxygen, nitrogen, and so on, can assemble themselves into orderly molecular structures, and larger molecules, such as those of tropocollagen, or protein subunits in general, can assemble themselves into complexes whose structure is dependent on localized and directional intermolecular forces. It seems that such comparatively large entities as the units that come together to form the head structures of bacteriophages or bacterial flagella are capable of orderly self-assembly, but the chemical forces that give rise to the interunit bonds are still little understood.
Processes that fall into the same general category as self-assembly may occur within aggregates of cells. The units that self-assemble are the cells themselves. Interaction and aggregation may be allowed to occur in assemblages of cells of one or more different kinds. In such cases it is commonly found that the originally isolated cells tend to adhere to one another, at first more or less at random and independently of their character, but later they become rearranged into a number of regions consisting of cells of a single kind. When the cells in the initial collection differ in two different characteristics, for instance in species and organ of origin, the assortment in some cases brings together cells from the same organ, in other cases cells from the same species. Mixtures of chick and mouse cells, for instance, reassort themselves into groups derived from the same organ, whereas cells from two different species of amphibia sort out into groups from the same species more or less independently of organ type.
This morphogenetic process probably has only a restricted application to the formation of structures in normal development, in which only in a few tissues (e.g., the connective system) do cells ever pass through a free stage in which they are not in intimate contact with other cells, and cells of different origin do not normally become intermingled so as to call for processes of reassortment. To explain normal morphogenetic processes of plants and animals one must look to the results that can be produced by the differential behaviour of cells that remain in constant close contact with one another. Several authors have shown how striking morphogenetic changes could be produced within a mass of cells that remain in contact, but that undergo changes in the intensity of adhesion between neighbouring cells, in the area of surface in the proportion to cell volume, and so on. |
|
|
|
|
Differentiation
Differentiation (B)
Differentiation is simply the process of becoming different. If, in connection with biological development, morphogenesis is set aside as a component for separate consideration, there are two distinct types of differentiation. In the first type, a part of a developing system will change in character as time passes; for instance, a part of the mesoderm, starting as embryonic cells with little internal features, gradually develops striated myofilaments, and with a lapse of time develops into a fully formed muscle fibre. In the second type, space rather than time is involved; for instance, other cells within the same mass of embryonic mesoderm may start to lay down an external matrix around them and eventually develop into cartilage. In development, differentiation in time involves the production of the characteristic features of the adult tissues, and is referred to as histogenesis. Differentiation in space involves an initially similar (homogeneous) mass of tissue becoming separated into different regions and is referred to as regionalization.
Histogenesis involves the synthesis of a number of new protein species according to an appropriate timetable. The most easily characterized are those proteins formed in a relatively late stage of histogenesis, such as myosin and actin in muscle cells. The synthesis of proteins is under the control of genes, and the problem of histogenesis essentially reduces to that of the genetic mechanisms that direct protein synthesis.
Regionalization is concerned with the appearance of differences between various parts of what is at first a homogeneous, or nearly homogeneous, mass. It is a prelude to histogenesis, which then proceeds in various directions in the different regions so demarcated. The processes by which the different regions acquire distinct contrasting characteristics must be related to some of the processes discussed under morphogenesis. Unlike morphogenesis, regionalization need not involve any change in the overall spatial shape of the tissues undergoing it. Regionalization falls rather into the type of process for which field theories have been invoked. |
|
|
|
|
| |
Control And Integration Of Development
|
Control And Integration Of Development
Control And Integration Of Development (B)
No text. |
|
|
|
Phenomenological aspects
Phenomenological aspects (B)
One of the most striking characteristics of all developmental systems is a tendency to produce a normal end result in spite of injuries or abnormalities that may have affected the system in earlier stages. In many cases, perhaps in most, only injuries inflicted during a certain restricted period of development can be fully compensated for. During such periods the system is said to be capable of regulation or the restoration of normality.
Developmental regulation is often discussed in terms of homeostasis, or regulatory mechanisms. Many systems, including biological ones, exhibit a tendency to return to initial equilibrium once they are diverted from it. A developing system is, by definition, always changing in time, moving along some defined time trajectory, from an initial stage, such as a fertilized egg, through various larval stages to adulthood, and finally to senescence. The regulation that occurs in such systems is a regulation not back to an initial stable equilibrium, as in homeostasis, but to some future stretch of the time trajectory. The appropriate word to describe this process is homeorhesis, which means the restoration of a flow.
A second major phenomenological characteristic of development is that the end state attained is not unitary but can be analyzed into a number of different organs and tissues. The overall time trajectory of this system can, therefore, also be analyzed into a number of component trajectories, each leading to one or another of the end products that can be distinguished in the later stages. A major discovery of the early experiments on developing systems was that, in many cases at least, the different time trajectories diverge from one another relatively suddenly during some short period of development, which usually occurs well before any visible signs of divergence can be seen microscopically or by any other available means of analysis. The most dramatic and influential example of this was provided by studies on the development of the amphibian egg at the time of gastrulation, or formation of a hollow ball of cells. At this time the lower hemisphere of the embryo will be pushed inward (invaginated) to develop into the mesoderm and endoderm, and the upper hemisphere will remain on the surface, expanding in area to cover the whole embryo. Approximately one-third of the upper hemisphere will develop into the nervous system and the remainder into the skin. During the period when these morphogenetic movements of invagination and expansion are occurring, a process takes place by which a portion of the upper hemisphere enters a trajectory toward neural tissue and another part enters a trajectory leading to epidermal development. This process of determination of developmental pathways happens relatively quickly, during a period when the cells of the two different regions appear superficially alike. The occurrence of the determination can in fact be demonstrated only experimentally. Before it occurs, any part of the hemisphere can develop either into neural tissure or into skin. After it has happened, each part can develop only into one or the other of these alternatives.
It is clear that an adequate theory of development has to account not only for the processes by which a developing system moves along its appropriate time trajectory, but also for the nature of the processes by which the trajectories diverge from one another and become fixed or determined in the developing cells.
The determined state can be transmitted through many cell generations. An example of this transmission can be seen in Drosophila flies. The imaginal buds of Drosophila are small packets of cells that become separated from the main body of the embryo in the early stages of development. They persist throughout larval life and then enter into the differentiation of adult characteristics when stimulated to do so by the hormones secreted at the time of pupation. These pupation hormones disappear from the body of the adult insect, and imaginal buds transplanted into the body cavity of an adult undergo many cell generations, but they do not show any signs of differentiating into the specific tissues of the corresponding adult organ. After many generations of proliferation, however, the cells can be transplanted back into a larva ready to pupate; they thus submit to the pupation hormones and differentiation occurs. Through many generations of proliferation the cells have retained the determination as to which adult organ they will develop into when the pupation hormones become available.
Attempts to identify the determining agent have not yet been successful. Experiments on amphibian eggs, however, have given rise to one important general conclusion; namely, that the process of determination can take place only during a certain period of development, in which the cells of the upper half of the amphibian egg are poised between the two alternatives of development into neural tissue or into skin. They are said at this time to be “competent” for one or the other of these types of development. While they are in this state, and only while they are in it, a variety of external agents can switch them into one or the other of the possible pathways. Such a situation may be contrasted with one in which the cells were neutral, or featureless, and required then an external agent to transmit to them the quality of becoming nervous tissue or of becoming skin. This would mean that the reacting cells required information or instructions to be added to them from outside. Such a situation is not characteristic of biological development. Both in highly developed organisms such as amphibians and in simpler ones such as bacteria, the external agents act only as a releaser that switches on one or another process for which all of the necessary information is already incorporated in the cells concerned. |
|
|
|
|
Analytical aspects
Analytical aspects (B)
The existence of these developmental phenomena was realized in the first third of this century. During this period, biologists had no clear notion of the fundamental concepts needed to explain development. Developmental biologists, or embryologists, attempted to account for their observations by means of ill-defined notions, such as “potencies” or “organ-forming substances,” or by referring to cellular properties that are real enough but obviously in themselves complex and essentially secondary in nature, such as cellular adhesiveness, the capacity of cell surfaces to differentially absorb certain substances, and so on. It was only gradually that developmental biologists came to realize the importance of the demonstration by genetics that nearly all the instructions required for the building of a new organism are contained in the genes that come together during fertilization, and that the small additional amount of information, contained primarily in the ovum, is itself a product of genetic instructions provided in the body of the mother in which the ovum is produced. The fundamental problems of the theory of development are, therefore, to understand how these units interact with one another to form more complex mechanisms that bring about the cellular or tissue behaviours of the different types of developing systems.
In the development of the neural system of vertebrates, for example, a great many genes must be active in controlling the synthesis of particular proteins. In the formation of the wing of a Drosophila, the activity of some 20 or 30 genes has been definitely demonstrated, and certainly many more are involved. The action of all these genes, however, must be considered to form a network involving many types of feedback and other interactive loops, the overall result of which is a product in which many components are present in precisely defined concentrations; and further, the developmental process leading to this end result must be buffered or stabilized, in the sense that if the process is diverted from its normal course at an early stage, it returns to some later stage of the normal trajectory. The realization that the basic units of development are genes indicates that a stabilized time trajectory involves the action of tens, if not hundreds, of genes. The realization that biological development is fundamentally an expression of the controlled activities of genes has finally resolved one of the old philosophical controversies about the nature of development, between preformation and epigenesis. The former supposed that, at the initiation of development, for instance in the fertilized egg, the system already contained some representative of every organ that would eventually put in an appearance. The vindicated theory of epigenesis, on the other hand, supposed that later appearing entities were produced during the course of development.
The modern interpretation of epigenesis is that the initial stage of development does contain certain entities with well-defined properties, namely the genes. These do not, however, represent directly the later formed organs, which arise by the gradual interaction and progressive unfolding of the properties of groups of genes.
One of the major problems confronting modern developmental biology—namely, the nature of “determination”—requires an understanding of how genes are “primed” to enter into activity when an appropriate stimulus is given. The state of priming presumably has to apply to quite a large number of genes, though perhaps not to all that will be involved in the stabilized, or buffered, time trajectory, since some may be brought into activity by the operation of the earlier active ones. The priming, moreover, has to be able to persist through cell division and be capable of transmission through many generations of cell proliferation. Few concrete suggestions as to the mechanism have yet been made. One is that the primed genes are already producing the ribonucleic acid molecules, called messenger RNA’s, which direct protein synthesis in the cell, but that these messengers are in some way inactivated or prevented from activating the protein-synthesizing machinery; this is known as the “masked messenger” hypothesis. Arguments in favour of this hypothesis are, however, circumstantial rather than direct. In some cases, for instance that of the Drosophila imaginal buds, there is direct evidence against it. Another hypothesis, perhaps more attractive, but much vaguer, is that the determination or priming involves the intervention of some of the large amounts of reiterated DNA known to be present in the cells of higher organisms. At the present time, however, biology lacks any convincing theory of determination in terms of gene action.
It appears at first sight that more is known about actual differentiation than initial determination. Actual differentiation must involve the controlled synthesis of particular proteins, coded for by specific genes. Certainly, a great deal is known about the mechanisms that control the action of genes in directing the synthesis of proteins in simple organisms such as viruses and bacteria. It is tempting to suppose that similar systems operate in controlling the synthetic activities of genes in higher organisms. Unfortunately, no single case of an exactly similar controlling system has ever been discovered in higher organisms, in spite of an intense search for it. It may in fact be suggested that until there is a fuller understanding of the mechanism of “priming” genes at the time of determination, there can scarcely be an adequate account of the way in which the activity of these genes is controlled at later stages. |
|
|
|
|
| |
Development And Evolution
|
Development And Evolution
Development And Evolution (B)
Evolution is carried out by a process dependent on mutation and natural selection. Expositions of this thesis, however, tend to overlook the fact that mutation occurs in the genotype, whereas natural selection acts only on the phenotype, the organism produced. It follows from this that the theory of evolution requires as one of its essential parts a consideration of the developmental or epigenetic processes by which the genotype becomes translated into the phenotype. The consequences of such considerations are discussed in the following sections. |
|
|
|
Effect on life histories
Effect on life histories (B)
No text. |
|
|
|
Length and timing of the reproductive phase
Length and timing of the reproductive phase (B)
Natural selection results in the production by one generation of offspring that are able to survive and reproduce themselves to form a further generation. The time unit appropriate to natural selection is therefore the generation interval. There will always be some natural selective pressure for the shortening of the generation interval, simply out of a natural economy, and for an increase of the number of offspring produced by any reproducing individual. One of the ways in which such an increase could be assured would be the lengthening of the reproductive phase in the life history; another would be an increase in the number of offspring produced.
These are, of course, not the only natural selective pressures that operate. It is clear enough that, in evolution, they have often been overcome by other pressures. There is another natural selective pressure of more general importance. This is the pressure to restrict the length of the reproductive period, and indeed to remove reproductive individuals, in order to make room for the maturation of a new generation in which new genetic combinations can be tried out for their fitness. A species whose individuals were immortal would exhaust its possibilities for future evolution as soon as its numbers saturated all the ecological niches suitable for its way of life. Death is a necessary condition for the trying out of new genetic combinations in later generations. It is usually brought about, in great part at least, by combinations of two processes: restriction of the period of effective reproduction to a certain portion of a life history, and as a necessary consequence of this, the absence of natural selection for genetic mutations that would be effective in preserving life after reproduction has ceased. In some organisms—for instance, long-lived trees—there may be no restriction of reproduction to a particular period in the life history, but their development involves the gradual accumulation of larger and larger quantities of nonliving materials, such as dead wood, which presents a growing handicap, in the face of which the organism cannot indefinitely maintain itself against the inevitable hazards of existence. It is still something of a question whether these natural selective forces are sufficient in themselves to account for the phenomena of senescence, aging, and eventual death, which are found in various forms throughout nearly the whole biological kingdom.
As was mentioned above, evolution has produced a number of the types of multiphasic development, in which the life history involves a succession of larval stages. Such types of development offer the possibility of changing the relative importance of the various stages in relation to the exploitation of resources and reproduction by the species. There are, for instance, many types of animals (particularly insects) in which nearly the whole life history is passed in a larval stage in which most of the feeding and growth of the organism is carried out, the final adult stage being short and used almost entirely for reproduction. Another evolutionary strategy has been to transfer the reproductive phase from the final stage of the life history to some earlier larval stage. This again has occurred in certain insects. If such a process is carried to its logical evolutionary conclusion, the final previously adult stage of the life history may totally disappear, the larval stage of the earlier evolutionary form becoming the adult stage of the later derivative of it. An example in which this process is at least partially accomplished is in the axolotl, a salamander that reproduces in a larval stage and in nature rarely if ever metamorphoses into the adult, but can be persuaded to do so if injected with extra supplies of the hormone thyroxin. It has been suggested that such processes of neoteny (the retention of some juvenile characteristics in adulthood) have played a decisive role in certain earlier phases of evolution, evidence of which is now lost. It has been argued that the whole vertebrate phylum may have originated from modifications of one of the larval stages of an invertebrate group. |
|
|
|
|
Recapitulation of ancestral stages
Recapitulation of ancestral stages (B)
The modifications of life histories just mentioned are aspects of a more general situation; namely, that the only variations that can become available for natural selection to operate on are those that can be produced by alterations of the developmental or epigenetic system of an existing organism. Any new mutant gene can cause a change only in a preexisting set of developmental interactions; the phenotypes to which it can give rise are limited by the nature of the system that it will modify. One immediate result of this situation is that the development of a later evolved form will retain many features from the development of its ancestors: most evolutionary developments are likely to be additions to the previous organization. Since there is evolutionary pressure to reduce the length of time between generations, the addition of a new feature to development is likely to be accompanied by a speeding up of the older stages, and probably omission of certain of them.
To repeat, the development of a late-evolved form retains those aspects of earlier life histories that are essential for the building up of later developmental stages that may be important for natural selection. In the vertebrates, for instance, highly evolved types such as mammals and birds produce during their early development remnants of the primitive kidneys (pronephros and mesonephros) that functioned as excretory organs in their evolutionary ancestors. Although these organs no longer perform their physiological functions in later organisms, they play an essential role during the formative processes of embryonic development. Some structures characteristic of evolutionary ancestors may be retained for relatively short evolutionary periods after they have lost their original function simply because there is not sufficient natural selective pressure to bring about their elimination when they no longer have any obvious function, either physiologically or epigenetically; the human appendix is an example. |
|
|
|
|
Adaptability and the canalization of development
Adaptability and the canalization of development (B)
A developing organism is subjected to natural selection by its particular environment. The environment is not the same for all individuals of a population, nor does it necessarily remain the same throughout evolutionary periods of time. An organism can be regarded as having to meet environmental changes that are unpredictable. There are basically two different types of strategy employed, in various proportions in different organisms, to meet this situation. One, perhaps the more obvious, is to evolve a high capacity for modification by environmental circumstances in ways that increase fitness in the environment in question; this is the strategy of increasing adaptability. It is probably true to say that all organisms show some capacity for adaptation, either short-term (physiological) or longer term (developmental), to their environments. In most organisms, however, particularly in most higher organisms, there is considerable development of the alternative strategy, which is to build up well-buffered or channelled developmental processes, which lead to the production of a relatively predictable invariant end result in the face of very diverse environments. The second strategy is likely to be followed in situations in which the environment is likely to change markedly during the course of the organism’s life.
Whether or not this is the main reason for the evolution of channelled, or canalized, developmental systems, a considerable degree of canalization is very common. It is relatively rare to find instances in which the form of an animal is highly dependent on the early environment, although such dependence is common enough among plants. Much more frequently, situations such as that typified by the house mouse are encountered: the mouse develops into an almost identical form whether it lives in the tropics or in a cold-storage depot.
This canalization of development severely restricts the phenotypic effects that can be produced by mutations. In particular, many new mutations occurring in a single dose in a diploid organism are found to be recessive, or ineffective in causing any alteration in the phenotype. As this discussion makes clear, canalization should not be considered as a relation involving only the normal and mutated forms of a particular gene, but rather the result of the interaction of many genes. |
|
|
|
|
Genetic assimilation
Genetic assimilation (B)
A long-standing controversy in biology has been concerned with whether phenotypic modifications produced by abnormal environments are heritable in the sense that they can be produced by later generations in the absence of the original environmental stress. The hypothesis that they are heritable was advanced by the French evolutionist Lamarck in the 18th century and is generally known as the “inheritance of acquired characters.” It found some supporters among biologists, some of whom used it as an argument against the Darwinian theory of evolution. In a broad sense, all characters are to some extent inherited, in that they depend on the genotype of the organism, and to some extent acquired, since development is also affected by the environment. In a stricter sense, however, Lamarck’s hypothesis suggests that there is some inherent biological property that enables organisms to pass on physical modifications to their descendants, independently of a Darwinian mechanism of selection.
The combination of adaptability and canalization in development can explain such phenomena in strictly Darwinian rather than Lamarckian terms. The abnormal environment acting during development may succeed in modifying even a well-canalized development system. If the modification is of an adaptive kind and increases the fitness of the individuals in the unusual environment, it will be favoured by natural selection. The development of the selected individuals will, however, also show some properties of canalization, that is to say, resistance to further environmental changes. This invariance may be sufficient to prevent offspring of the selected individuals from reverting completely to the original phenotype even if they are removed from the abnormal environment. After selection for an adaptive modification in an abnormal environment has proceeded for many generations, a form may be produced whose canalization is strong enough to maintain the new phenotype almost unaltered when the environment reverts to what it was before the abnormality occurred. This process, which has been demonstrated in a number of laboratory experiments, is known as genetic assimilation. It produces exactly the same results as those emphasized by advocates of the Lamarckian inheritance of acquired characters, but it produces them by an orthodox Darwinian mechanism operating on developmental systems that have the common properties of canalization and adaptability. It provides the most convincing explanation for the evolution of organisms that are physiologically or functionally adapted to the demands their way of life will make. |
|
|
|
|
|
|
|
|
|
|